Music from Plant Biosignals: A Conceptual and Analytical Orientation
Paul V. Miller and Christopher Cox
KEYWORDS: electronic music, posthumanism, phytosemiotics, sound art, ambient, postminimalism, bioelectricity, Mamoru Fujieda, Augustine Leudar, Data Garden, algorithmic composition
ABSTRACT: This paper provides a broad but comprehensive foundation for understanding the widespread practice of using the bioelectric properties of plants to create music and sound. We begin by describing how plant biosignals are harvested by several commercial electronic devices and a custom-designed interface. Then, we analyze large data samples of potential musical content generated by such technologies. Next we investigate the artistic and musical choices artists make when they interpret this data, analyzing work by British sonic artist Augustine Leudar, the Philadelphia Museum of Art’s Data Garden Quartet, and Japanese composer Mamoru Fujieda. Finally we position the entire enterprise within a posthumanist semiotic framework, specifically referencing work by Uexküll, Deleuze, Krampen, Watkins, and Marder. This interpretation allows us to speculate on the nature of communication and musical collaboration that can potentially take place between the human and vegetative domains.
DOI: 10.30535/mto.30.1.6
Copyright © 2024 Society for Music Theory
1. Introduction
[1.1] The practice of using plant bioelectric signals in sound installations and musical art has exploded in recent years. Its interdisciplinary nature has made analysis challenging because “plant music” lies at an intersection of music theory, biology, electrical engineering, computer music, aesthetics, and metaphysics. In this paper we examine the practice—which we term “electronic plant music” (EPM) and “electronic plant sound” (EPS)—from a holistic, semiotic perspective. We understand EPM and EPS to mean music or sound art whose macro- or microstructure is related to, or in some way generated by, plant-derived bioelectric signals.(1) We describe how bioelectric signals arise in plants, show how they are typically acquired and processed, and analyze several large biodata samples in a way that sheds light on creative processes. We then analyze three significant EPM/S projects by Augustine Leudar, the Data Garden Quartet, and Mamoru Fujieda. The results of our musical analysis cascade into an emergent critique guided by writings of Rupert Till and Kyle Gann. We see a correlation between the amount of intervention an artist makes interpreting biosignals and the work’s stylistic affinities to ambient and postminimalist music. We conclude by revisiting the field of phytosemiotics using critical posthumanist frameworks suggested by Uexküll, Deleuze, Krampen, Watkins, Marder and others. This article provides a provocative and comprehensive foundation for future work in this evolving area.
[1.2] We believe that locating EPM/S within the area of music theory provides particular advantages, principally because the main communicative realm and ultimate purpose of most projects in this field lies in the auditory domain. Music theory offers a variety of tools that reveal information about creative processes, musical structures, and listeners’ responses. Nevertheless, it is prudent to embrace a holistic, “big tent” approach to understanding the phenomenon of EPM/S due to the novelty of the enterprise. We are guided by Kofi Agawu’s sage advice put forth many years ago: “. . . given the relative paucity of analyses, erecting barriers against one or another approaches seems premature” (Agawu 1987, 196). In this spirit our essay casts a wide net, but we emphasize that the music-theoretical methods we deploy do not proscribe applying other critical methodologies to the domain of EPM/S, particularly those that privilege extramusical domains.
[1.3] The semiotics of Jean-Jacques Nattiez (1990) guides our holistic discourse. By understanding musical works as “symbolic forms,”(2) Nattiez provides strategies to “grasp the phenomena associated with the musical fact in relation to the materiality of musical works” (Nattiez 1990, 95), an objective that seems well-suited to our pursuit. Following Nattiez, we divide a musical work’s ontology into three levels or “registers”: the poietic (or creative) level; the neutral level, comprising the “trace” or written-out “score” (if there is one); and the esthesic level, which encompasses the audience’s response. Because the network of relationships between levels in EPM/S is quite complex, referring to these three registers brings methodological clarity.
Example 1. Nattiez: Six Analytical Situations from Music and Discourse: Towards a Semiology of Music (1990), 140
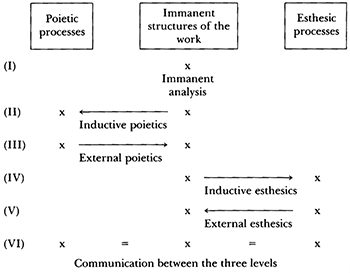
(click to enlarge)
[1.4] Nattiez’s “six analytical situations” together constitute what he calls a “geography of analysis” (Nattiez 1990, 140). These situations catalog analytical praxes or relationships between the three registers, clarifying the epistemological claims an analysis makes (Example 1). Thinking in this way also proves advantageous to our study, since certain aspects of EPM/S are frustratingly inaccessible; for example, it is not always possible to know what a plant responds to when it changes metabolic processes, because we cannot ask it, and it cannot tell us. Another potential stumbling block in EPM/S is that there is no guarantee of a material trace or score for the work, as many projects exist ephemerally “in real time.” However, Nattiez shows that it is possible to speculate about one level if we know something about another. This is a frequent situation we come across in EPM/S.
Example 2. Preliminary View of the “Analytical Situation” of EPM/S (after Nattiez 1990, 76)
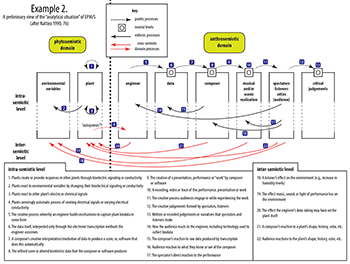
(click to enlarge)
[1.5] Nattiez wrote of the concept of the “analytical situation” more generally as a way to interpret analysis and better understand the claims it makes (Nattiez 1990, 135ff.). His example of the complex network of semiotic relationships in the Boulez/Chereau Ring cycle is one of the classics of modern music analysis (Nattiez 1990, 76). However, as intricate as the situation is with Wagner, it is even more daunting in EPM/S because the actors are not all human: a vexing anthrosemiotic/phytosemiotic divide must be crossed. Indeed, EPM/S could be understood as a quintessential posthumanist activity that invites us to reevaluate relationships between ourselves and the vegetal world. Following Nattiez, Example 2 captures our preliminary assessment of EPM/S’s peculiar and complex analytical situation. This example includes a brief summary of each relationship shown on the graph.
[1.6] While we cannot explore all the connections suggested in Example 2 within the scope of this article, we will provide a general overview of the most salient aspects. We start by analyzing the plant as a kind of autopoietic source of bioelectricity, then continue by scrutinizing techniques that engineers use to build systems that transcribe bioelectric data. Next we consider the structural characteristics of this “raw” material by examining data from several devices built specifically for EPM/S applications. We continue by analyzing various poietic and esthesic processes composers (or software) use to curate these data, which becomes material ready to create sound or music. We then shift our attention by turning towards actual musical works, focusing on three substantial but very different EPM/S projects. This analysis leads us to hypothesize that the way artists interpret biodata tends towards certain musical genres or styles. We conclude by plunging into metaphysical and ethical questions surrounding EPM/S, particularly its role in posthumanism thought.
2. Bioelectricity
[2.1] By our definition, the process of EPM/S begins with electrochemical reactions inside a living plant. In contrast to conventional musical instruments, which make sound by converting mechanical energy into vibrational (pressure) waves, EPM/S is created by converting a plant’s electrical properties into sound pressure waves via electronic technologies. Unless plants are moved by external forces such as the wind, animals or other external actors, they do not create sounds that are audible by humans.(3) However, it has been known for well over a century that plants are electrically active organisms (Darwin 1898; Burdon-Sanderson 1873). Plants continually produce electrical signals, which they transmit through their tissues. They generate electric potential by transporting charged ions (electrolytes) from cell to cell. Gates in cellular membranes regulate this ionic flow (Canales, Henriquez-Valencia, and Brauchi 2018). Possessing no specialized cells, neurotransmitters, or synapses to facilitate electrical conductivity, signals propagate through extracellular tissue more slowly than in animals.(4) Electrical signals regulate many processes crucial to plants’ survival, including respiration, protein synthesis, gravitropism, and leaf movement (Fromm and Lautner 2006). Biologists have cataloged three distinct classes of signals: action potential (AP), variation potential (VP) and system potential (SP) (Stankovic, Witters, Zawadski, and Davies 1998, Xie, Wu, Xing, Li, Chen, Duan, and Zhu 2021, Johns, Hagihara, Toyoto, and Gilroy 2021).
[2.2] While AP, VP and SP are typically transmitted through the xylem and phloem tubes in plant stems, research has shown that the electrical characteristics of plant leaves are predictable, cyclic, and correlated to an endogenous 24-hour circadian clock. Studies of two plant species demonstrated that the fundamental electrical characteristics of resistance, capacitance, and impedance change cyclically throughout the day (Xie, Wu, Xing, Li, Chen, Duan, and Zhu 2021). The variability is not particularly complex.(5) Leaf conductivity corresponds to the amount of water contained within it, which varies according to a plant’s rate of respiration throughout the day and night. There is considerable difference in leaves’ electrical conductivity between various plant species (Duursma et al. 2019).
[2.3] Because plants transmit electrical signals does not mean that they have a “consciousness,” though the constitution of plants’ perceptive worlds is an area of controversy and will be discussed later.(6) The scientific community initially reacted strongly (and somewhat uncharitably) against claims arising from uncontrolled experiments that plants possessed some form of consciousness (Tompkins and Bird 1973; Galston and Slayman 1979; Pollan 2013), but recent laboratory-based studies have yielded a more nuanced view (Shabala and Mancuso 2007; Baluška, Mancuso, and Volkman 2006, 2009, Affifi 2013). Established in 2005, the International Laboratory of Plant Neurobiology at the University of Florence (Italy) is a major center for research in this area.(7) The new scientific consensus is that plants have robust abilities to sense and react to conditions around them and can even learn and remember certain behaviors. This paradigm has inspired many EPM/S artists.
3. Harvesting bioelectric signals
[3.1] In a laboratory setting, AP, VP and SP signals are measured by inserting small synthetic or organic probes directly into inter- or intracellular spaces (Fromm and Lautner 2006; Macedo et al. 2021). However, artists will not typically have access to such elaborate apparatus. Instead, most use a non-invasive method derived from a principle first applied to so-called “lie detector tests”: galvanic conductance. Galvanic conductance is the amount of electrical conductivity between two electrodes across some substrate, such as skin or a leaf. The first to popularize this method in plants was Cleve Backster (1924–2013), a career CIA interrogation specialist whose highly questionable experiments in the 1960s involved attaching a polygraph instrument to plant leaves (Tompkins and Bird 1973; Galston and Slayman 1979). The electrodes employed in most EPM/S applications are called TENS (“Transcutaneous Electrical Nerve Stimulation”) cables, which were not originally intended for use on plants but work well nevertheless (Larimore 1984).
Example 3. Typical EPM/S setup showing TENS cables positioned approximately 4cm apart on the leaf of a croton houseplant (this was the methodology used throughout this study)

(click to enlarge)
[3.2] In a typical musical or sound application, a set of TENS electrodes are attached to a plant leaf. They adhere using an electrically conductive gel, which damages the leaf if it is not rigid enough. Consequently, plants with relatively strong, fibrous leaves are typically employed in EPM/S applications.(8) Electrically conducting clips are one alternative to TENS cables, and other applications use a clip and a probe that interfaces with a plant’s root system. The length of the cable connecting the electrode to the electronics needs to be short; otherwise, the faint electrical signal can be degraded by noise. Whatever their design, electrodes typically fulfill four requirements: they are robust enough to maintain contact with the leaf surface; they should not damage the leaf; they minimize electrical noise; and they must be electrically conductive. Example 3 shows a typical TENS electrode setup.
[3.3] The poietic role of the engineer includes building sensors, hardware, and software systems that transcribe plant biosignals into some sort of information usable on a computer or in a synthesizer. Engineers have devised many ways to do this, but in most implementations it is unclear to audiences and artists how musical content is generated. Some composers and sound designers have stepped into the engineering role and designed their own technologies, but this is the exception. Shedding light on this step empowers creators with agency over their material and suggests ways that new systems can be devised that are better tailored for specific aesthetic goals. We argue that the engineering step affects the final sonic outcome of EPM/S in many significant ways. In the next section we scrutinize several systems that produce content that musicians or sound artists might use.
Example 4. Six EPM/S devices/instruments

(click to enlarge)
[3.4] Once electrodes have been put into place, electrical biosignals can be converted into some kind of information artists can use. In most cases, this is MIDI data. Adopted in 1983 and improved in 2020, MIDI is a widely adopted digital musical standard that ensures compatibility among electronic musical instruments. Essentially, MIDI is a seven-bit integer (ranging from 0–127) that denotes pitch (middle C = 60), with a corresponding integer indicating velocity (loudness).(9) However, MIDI is only one of many ways that biodata can be transcribed. The next subsection analyzes six varying techniques of creating biodata usable to musicians, noting their structural pathways, and evaluating their advantages and shortcomings.(10) The six devices—or “instruments”—are shown in Example 4.
MIDISprout
[3.5] Released commercially in 2014 by a company called Data Garden, MIDISprout was the first widespread commercial biosonification device available to EPM/S creators. Its enormous success was due to the fact that it was simple to use, relatively inexpensive, and produced easy-to-manipulate MIDI data. Its inventor, Sam Cusumano, publicly released MIDISprout’s source code along with plans for the printed circuit board (PCB) and files for 3D printing the enclosure.(11) The MIDISprout runs a small electrical current through one TENS electrode and detects the electrical signal through the leaf from the other one.(12) This low-level voltage oscillates a 555 timing chip that generates electrical square waves. The duration of the high voltage pulse in the 555’s square wave output corresponds to the amount of galvanic conductance in the leaf. The MIDISprout system measures the duration of this “high” pulse, buffers 10 samples, shaves off the first and last sample, and computes the standard deviation of the remaining 8 samples.(13) Finally, the algorithm compares the standard deviation of the sample buffer to a threshold value set by a dial on the device itself. If the standard deviation is above the threshold value, MIDISprout generates a MIDI note scaled to the high and low values of the 8-sample data buffer. Consequently, MIDI notes are not related to the amount of conductance in a leaf itself, but rather to minute changes in electrical conductance over brief slices of time.(14) Sound or music is produced by connecting the MIDISprout into a DAW (Digital Audio Workstation) using a 5-pin MIDI cable.
PlantWave
[3.6] The MIDISprout’s success prompted Data Garden to release a second-generation device in late 2022 called PlantWave. The architecture of PlantWave is similar to MIDISprout, but with one important difference: PlantWave can sonify MIDI data right “out of the box” through a software synthesizer on a downloadable smartphone app. Users select different synthesizer patches with evocative names derived partly from Buddhist meditation practices. Music then plays through the smartphone. PlantWave can also transmit MIDI data through a USB-C cable or WiFi network.
DIY MIDISprout kit
[3.7] MIDISprout creator Sam Cusumano also offers a do-it-yourself (“DIY”) MIDISprout kit online. This device has capabilities comparable to PlantWave. Collecting various commercially available components, the kit consists of a small motherboard containing a 555 timing chip, colorful LEDs, and interface jacks coupled to a modular “AdaFruit EPS32 Featherboard.” The AdaFruit board connects to the motherboard by a detachable bridge so that Featherboards can be swapped in and out for different functionality as desired.(15) The device can transmit MIDI wirelessly like PlantWave, or through a UCB-C cable. The kit runs Cusumano’s open-source software, which comes preloaded on the device’s microcontroller.
Damanhour’s “Music of the Plants”
[3.8] A religious community of social activists in Italy called the Damanhour Foundation produces a device called “Music of the Plants.”(16) The instrument is a lightweight bamboo box about the size of a smartphone. The Damanhour electrode consists of a small clamp and a large ground pin that connect to the box. It includes an onboard synthesizer and its own internal speaker.(17) The Damanhour instrument offers several different preloaded patches that, like PlantWave, are named for objects in nature or Buddhist meditation practices. The device is particular: if it does not sense a strong enough biosignal, it will not produce output, notifying the user on its LCD screen that there is a “bad connection.” In addition to producing EPM right out of the box, Damanhour can send MIDI data to a DAW.
Custom design
Example 5. A custom passive amplifier design for plant bioelectric signals
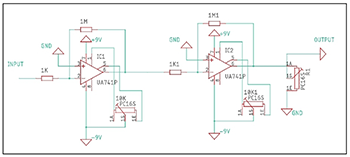
(click to enlarge)
[3.9] It is possible to construct low-cost interfaces from off-the-shelf electronic components. We fabricated a passive amplifier using two high-impedance op-amps mounted to a custom-etched circuit board and sent the output signal to a LabJack U3-LV data acquisition device.(18) While the raw signal itself is usable, we processed it using the software Max/MSP. This is an inexpensive and effective methodology for some purposes. Example 5 shows a schematic diagram.
Scion
[3.10] The firm Instruo fabricates the “Scion” Eurorack module.(19) Instruo is owned by Jason H. J. Lim, who enjoys a significant following in the Eurorack world.(20) Advertised as a “Quad Random Voltage Generator / Biofeedback > CV” device, Scion has become ubiquitous in online videos of EPM/S performances. Part of its popularity lies in its integration with the Eurorack modular synthesizer format. Based on Cusumano’s MIDISprout source code, Scion provides features that make it easier to use within an existing modular synthesizer, including four CV (control voltage) outputs, attenuators, gate outputs, glide knobs, a built-in quantizer, and a sensitivity knob. Notably, Scion’s unmediated, “raw” 555 output jack allows direct access to the timing chip’s oscillations. The 555’s distinctive buzzy sound can be sampled at audio rate, but it can also be analyzed by software and used as input for generative algorithms.
[3.11] All of the commonly used active methods of gathering electrical signals from plants with TENS electrodes have one thing in common: they measure the electrical resistance in a leaf. It is important to understand that these data do not necessarily represent electrical signals generated by the plant itself, but rather reflect a more-or-less accurate measure of the water content in the leaf at a particular point in its daily respiratory cycle. The native bioelectrical signals that most biologists study—AP, VP and SP—can affect the rate of respiration, but TENS electrodes do not directly measure them. Future artists may be able to measure AP, VP and SP through more complex sensors. This would provide musicians with data that more intrinsically belongs to the plant itself, presumably liberating a more direct information pathway to the organism.
4. The Data: structural characteristics of bioelectric signals
[4.1] In this section we analyze eight large datasets we collected using systems described in section 3 for two purposes: first, to elucidate the kind of raw material that a typical EPM/S composer or sound designer might expect to receive, and second, to show that the engineering role is truly a poietic process that has a salient effect on the outcome by constraining and shaping it in various ways. The data presented in this section were acquired in an environment that EPM/S artists might typically encounter: an indoor room with windows across one wall.
MIDISprout Data
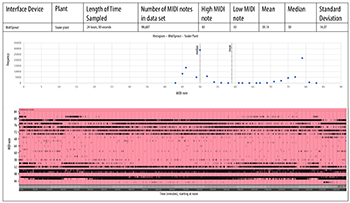
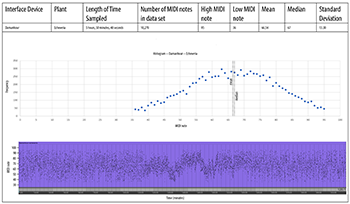
[4.2] We sampled the MIDI output generated by two plants of different species: Sansevieria trifasciata (snake plant) and Codiaeum (croton). TENS electrodes were placed on the same leaf about 4cm apart as shown in Example 3. Examples 6a and 6b show 24 hours of MIDI data continuously transcribed by the MIDISprout, starting at noon. Both examples show a “MIDI roll” at the bottom, which graphs the MIDI pitch (y-axis) over time (x-axis). Above, a histogram shows the number of times each MIDI note occurred in the data set. These data show that these two plants both produced slightly higher MIDI notes during the day than at night. However, the snake plant produced lower notes that were more widely dispersed, while the croton produced higher notes that were more compact.
Example 6a. MidiSprout, croton plant
(click to open PDF in new tab) | Example 6b. MidiSprout, snake plant
(click to open PDF in new tab) |
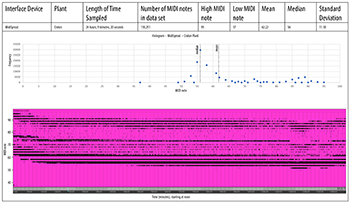
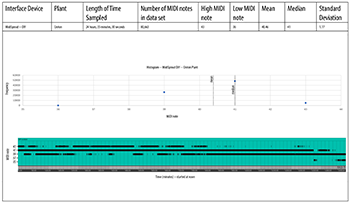
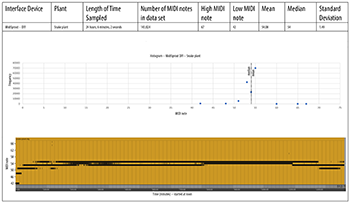
DIY MIDISprout Data
[4.3] We tested this device on the exact same plants as the MIDISprout, but the results (Examples 7a and 7b) were significantly different. Here, the MIDI pitch output is much more constrained than the original MIDISprout device, but notes tend to be higher for the snake plant, and lower for the croton. This is the reverse of the original MIDISprout data. The difference is probably because this device features an opto-isolator component that electrically insulates the 555 chip’s output signal as it runs into the Adafruit board, making the DIY MIDISprout less susceptible to signal noise.(21)
Example 7a. MidiSprout, DIY version, croton plant
(click to open PDF in new tab) | Example 7b. MidiSprout, DIY version, snake plant
(click to open PDF in new tab) |
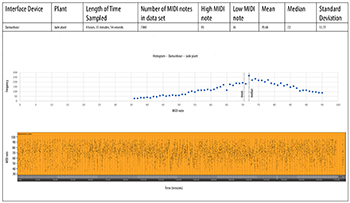
Dananhour Data
[4.4] We were limited to datasets of between 5 and 6 hours because of the internal battery’s capacity, and the inability to run the device on external power at the same time as it outputs MIDI data. We sampled a small jade plant and an echeveria succulent. These plants were used because the unusual root probe in the Damanhour electrode worked well with them. It was also a choice of necessity: the Damanhour will not output MIDI data if its software does not think it is receiving a strong signal. The data (Examples 8a and 8b) differed significantly from what we expected, probably because the Damanhour’s software prunes the raw data to produce EPM right “out of the box,” using its own built-in speaker.(22) Since the Damanhour schematics are not in the public domain, we could not examine them for this study. However, we noticed several important details. First, the system curiously tends to oscillate between producing MIDI notes between a fairly narrow range (what we might call “alto or soprano”) and a wider range of pitches. This effect is particularly noticeable in the jade plant data (Example 8a, in orange). If the composer or sound designer chooses to sonify this data with pitches, the musical effect will produce a sudden “opening up” in registral space that could create a pleasing outcome. We have no explanation for the very slow rising and falling arch in the middle of the echeveria data set (Example 8b, in purple, between 140 and 190 minutes), which was not observed elsewhere. Notably, the Damanhour device produced significantly higher MIDI notes on average than either MIDISprout.
Example 8a. Damanhour, jade plant
(click to open PDF in new tab) | Example 8b. Damanhour, echeveria plant
(click to open PDF in new tab) |
Custom Design Interface Data
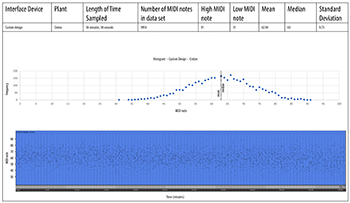
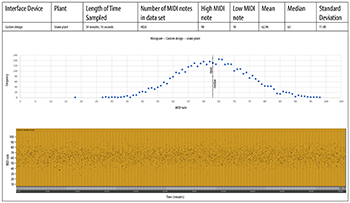
[4.5] We collected biodata from croton and snake plants simultaneously through two channels of our custom-designed amplifier system. A Max/MSP patch performed similar calculations on incoming data as Cusomano’s algorithm, but we modified some parameters to increase the variety of note durations. We used a slightly larger data buffer (10 samples) to compute standard deviation. Our passive amplifier operated without sending any voltage through the plant—and lacked a 555 timing chip—leaving ourselves open to a noisier biosignal. The entire system had to be operated on battery power, because plugging our laptop into a wall socket substantially increased the electronic noise. Although our datasets were each only about an hour long, we collected 7980 MIDI notes for analysis. The data appears more randomly distributed than any other device, but the results (Examples 9a and 9b) could still be usable in some EPM/S applications. We noticed that the snake plant generated a significantly wider range of MIDI notes than the croton plant, suggesting that the system was in fact transcribing biosignals that were structurally different from plant to plant. Both plants produced almost exactly the same average MIDI note (62.96 vs. 62.94). The advantage of our system was that it could transcribe several plant signals simultaneously due to the low cost of its electronic components. A future iteration of this device could include better noise isolation hardware.
Example 9a. Custom passive amplifier, croton plant
(click to open PDF in new tab) | Example 9b. Custom passive amplifier, snake plant
(click to open PDF in new tab) |
Scion Data
Example 10. Scion Eurorack module: biodata transcription

(click to open PDF in new tab)
Example 11. Scion’s 555 timing chip (“raw output”): one second sampled at 44.1kHz, 24 bits
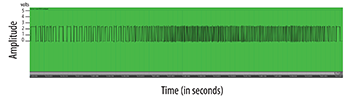
(click to open PDF in new tab)
[4.6] Although the Scion module is based on Cusumano’s algorithm, it produces control voltage instead of MIDI notes. Miller and Riordan collected useful preliminary data using the Scion instrument while subjecting test plants to various interventions including touching, burning and dramatic changes in sunlight (Miller and Riordan 2019). When we hooked our croton and snake plants up to Scion, we noticed the croton produced a small, constantly changing output signal, but the snake plant did not induce any variation whatsoever even when the sensitivity dial on Scion was turned to maximum.(23) Consequently we swapped the snake plant for an aloe plant to produce the data shown in purple colors in Example 10. This example shows two consecutive 50-minute datasets of Scion’s four CV outputs sampled at audio rate (44.1 kHz).(24) As can be seen most clearly in the aloe plant data (in cool colors), Scion’s four CV outputs progressively produce shorter segments of variable data, suggesting that each channel incrementally raises the user-defined threshold value in Cusumano’s algorithm. This would explain why the fourth channel in each dataset has the longest flat voltage output (flat lines). To augment our analysis of Scion’s capabilities we sampled its “raw” 555 output jack at audio rate to explore the structure of the square waves it produces. Example 11 shows 1 second of 555 output. This example vividly clarifies the raw signal that Cusumano’s algorithm processes.
[4.7] In the Scion datasets, our aloe plant produced dramatic variations in control voltage, separated by long stretches of static voltage. In some EPM/S applications, this could be a serious problem, because designers frequently look for CV that varies more continuously over time. However, the inherent flexibility of the Eurorack synthesizer workflow means that it is still possible to build a patch around the plant’s idiosyncrasies. For example, a sound designer might modulate a periodic control signal that generates a pitch ostinato by a signal like the one produced by the aloe plant. The effect might be an ostinato that repeats many times, only to be dramatically altered at unpredictable intervals. This example illustrates how important it is to analyze the transcribed biosignals beforehand, in order to build a patch that takes into account the plant’s own electrical profile and the module’s quirks.
[4.8] Our data show that the material artists have to work with varies dramatically according to the transcription method. The engineer’s role is itself a poietic aspect of the work because decisions made in this step significantly affect the choices available in the next creative stage. Mileece l’Anson—whose biosound projects have been featured at many prominent venues—writes of her sensor-derived material as a “live stream of variable data,” and a “continuous stochastic process” (l’Anson 2019, 502). However, l’Anson’s observation could be amended: a “useful” data stream ought to provide enough shape that an artist can create music or sound that has some variation, but not so much that it is just random noise. Balancing these two extremes is part of the esthesic work of the engineer, who responds to the idiosyncrasies inherent in plant biosignals to build a device that can supply the composer with an appropriate flow of usable data. The engineer thus fulfills both a poetic and esthesic role that goes beyond mere transcription.
5. Towards sound and music: three EPM/S projects
[5.1] An artist (or piece of software) must interpret plant biodata after it is transcribed from biological sources, transforming a stream of numbers such that they give rise to some immanent musical or sonic form. There are many ways to do this. In this section, we provide important context for this poietic process. Then we analyze three representative large-scale EPM/S projects that show the variety of possible artistic expressions.
[5.2] Data sonification is a term frequently applied to the process of transforming streams of data into auditory information. As discussed by Grond and Hermann (2012), data sonification employs one medium to represent something more than itself, and typically lays claim to “explanatory powers” by occupying a continuum somewhere between “mere illustration” and music. Perhaps the most straightforward representational strategy in EPM/S would be simply to map each MIDI value to a pitched note, resulting in a relatively unmitigated presentation of data. In this case, the main poietic decision the artist makes concerns the choice of instrument to represent the data. But as we have seen in the systems we have explored, these data frequently seem quite tonally unorganized. A common technique is to process MIDI data with a quantizer, which can map chromatic notes to diatonic scales (or any other desired pitch collection). Similarly, quantizers can operate in the rhythmic or time domain by moving note onsets and offsets to align with pulses in metric grids. Although EPM/S artists typically quantize pitch according to an equal-tempered scale, there is no inherent reason other tuning systems cannot be used instead. Devices such as the PlantWave, Damanhour, and Scion have built-in quantization options that provide little flexibility in this respect. DAWs present a much broader range of choices, and software such as Max/MSP or pd allow for considerably greater freedom.
Audio Example 1. Nightingale song, rhythmically quantized (samples ordered by biodata)
Audio Example 2. Nightingale song, not rhythmically quantized (samples ordered by biodata)
[5.3] More creative interventions are also possible. MIDI data can trigger changes in tempo, orchestration, or dynamics, affecting other musical parameters or even changing pitch quantization itself. The outcome can be understood less as data sonification and more as an “autonomous system” of automatic composition that follows an intricate set of rules. Even representing MIDI data as pitch is only one possibility among many: the same data can trigger indexed sound samples. One such implementation is our “synthetic birdsong.” First, software detects transients in a nightingale song, and then the song is sliced into short, indexed chunks— each beginning with a transient.(25) The plant biodata then sequences the slices. The result may either be time-quantized (Audio Example 1) or rhythmically free (Audio Example 2). Here, the relationship between the original plant-derived MIDI data and resultant sound is quite obscure.
[5.4] Many EPM/S artists encourage audiences to be aware that biological processes are somehow affecting the resultant sound, even if many layers of mediation remain opaque. Plants can become part of the performance not only by featuring their biosignals, but also through their visual display or exhibition. As we indicate in Example 2, it is possible to provoke an esthesic reaction in spectators not just to the sound, but also to the plant’s form, color, or smell. Miya Masaoka, whose “Pieces for Plants” performances involved displaying a lush array of ten or more houseplants, encouraged audiences to participate in actively affecting the sound that is produced. She wrote, “. . . people can approach the plant, and hear the plant’s response to their presence; as they move their hands around the plant at varying speeds, the plant’s responses are audible.”(26) While Masaoka encourages audience participation, others deter it. Although the projects we selected for analysis below only begin to represent the diverse practices of EPM/S, they range from what might be termed phytocentric (“let the plants speak for themselves”) to the anthropocentric (“let the audience or composer freely manipulate the plant and/or the biodata as desired”).
[5.5] Augustine Leudar’s tryptic of sound installations—entitled Surrounded, The Code, and El Bosque Encantado—were site-specific 3D environmental sound projects in the forests of Bolivia, Ecuador and Brazil.(27) Although Leudar specifically intended to avoid what he called “data sonification,” he took as his premise the idea that the pathways of electrical signals transmitted through a subterranean root and fungal network could be made audible.(28) Leudar intended to make “activity in this network tangible through sound in real time and immerse the public in the complexity of these processes” (Leudar 2018, 518), so his practice was clearly related to data sonification. As the basis of his sound construction, Leudar sampled ultrasonic sounds in the rainforest and pitch-shifted them downwards into human hearing range. He also created synthesized sounds, which were modified through various filtering, delay, reverberation, granularization and pitch-shifting techniques. His system actively panned these sounds around many loudspeakers that were placed next to trees’ biosensors. If a particular signal appeared in one tree and then in another, the system panned sounds along that pathway.(29) Leudar sonified an inferred underground bioelectric communication network that presumably operated in a small patch of forest. While he conceded that this work is still preliminary, Leudar’s installations were impressive demonstrations of sophisticated sound-processing techniques. They were also notable for his sensitivity to and incorporation of the rainforest’s native soundscape.
Example 12. Effects of touching a snake plant on biodata transcription (data recorded using custom passive amplifier)
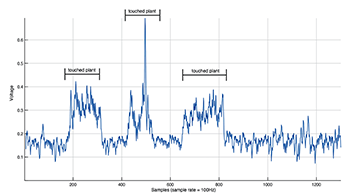
(click to enlarge)
[5.6] In contrast to Masaoka’s stance, Leudar’s aesthetic paradigm regarded plant “autonomy” as vital. He wrote:
In general it is best to avoid touching plants or even going near them . . . despite the seemingly dramatic response it provokes in electrode readings. . . . This is misleading and deprives the artist of an opportunity to present the true electrical processes happening within the plant. (Leudar 2014)
Touching plants does typically result in a rapid and dramatic electrical response, caused by the fact that humans both produce electric charge and can store it in our bodies. Example 12 illustrates how touching a plant leaf near a TENS electrode dramatically changes whatever signal the plant produces on its own. Leudar’s approach to EPS interpreted plant biodata in a way that attempted to elucidate its own internal structural characteristics. He drew a strict line between anthrosphere and phytosphere by discouraging physical contact between the two, heightening perceived boundaries.
Audio Example 3. Data Garden at the Philadelphia Museum of Art, from 14 April 2012 (20:15–20:56)
[5.7] The Data Garden Group employed a different strategy. Shortly before announcing the commercial production of the MIDISprout, three of their members—Sam Cusumano, Joe Patitucci, and Alex Tyson—produced a sound installation at the Philadelphia Museum of Art from April 13–15, 2012. Two hour-long tracks are available on Bandcamp and other online platforms.(30) Data Garden’s approach was to assemble an imaginary “bioband” of four distinct voices corresponding to invented musicians: lead synthesizer, rhythm, bass synthesizer, and ambient/effect pads. Data Garden’s playful approach lowered the stakes of their musical endeavor. Their sound installation layered dense collections of repeating diatonic and pentatonic notes over ambient pads. At times the textures varied, though the pitch patterns themselves were somewhat repetitive. Audio Example 3 gives a general sense of the first day’s soundscape.
Audio Example 4. Data Garden at the Philadelphia Museum of Art, from 15 April 2012 (43:2–43:53)
[5.8] In Data Garden’s music from the second day of their installation, occasional tempo changes in the lead (top) synthesizer part can be heard. This could have happened if their software used MIDI notes themselves from the plant interface to trigger such changes—a technique we suggested above in paragraph [5.3]. MIDI notes are not constrained to represent only pitches: a specific note (or sequence of notes) can be programmed to effectively change the tempo of a musical stratum, e.g. doubling or halving its speed. Whether or not Data Garden’s software worked in this way, several such changes are audible in the lead synthesizer part (Audio Example 4). Although this feature is prominent in the second day of Data Garden’s installation, both days’ music employs a constrained reservoir of diatonic or pentatonic pitches to generate slowly moving bass lines, and a larger set of diatonic pitches for more animated treble lines.
[5.9] While the Data Garden installation was innovative in many respects, the sonic result might strike some listeners as underwhelming or even redundant due to the high degree of repetition. Although structural changes in texture and rhythm are occasionally audible, the monotonic modality provides little variety on a larger timescale. Data Garden’s technique of using MIDI data to trigger changes in rhythm could be developed further in other musical domains. Even so, their overall accomplishment was impressive as the field was still quite new in 2012.
Example 13. Fujieda/Dogane, “Ecological Plantron,” liner notes
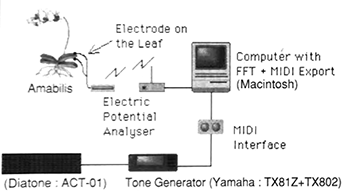
(click to enlarge)
[5.10] Composer Mamoru Fujieda (b. 1955) has worked with plants in his musical compositions for almost thirty years.(31) Fujieda’s initial experiment was in partnership with inventor and media artist Yuji Dogane (b. 1957). In 1994, the two produced an experimental album called “Ecological Plantron,” which, like Data Garden’s work, is an audio document of a sound installation that took place at a Tokyo art gallery.(32) Dogane’s “Plantron” was a device that converted bioelectric signals to MIDI notes using an FFT (Fast Fourier Transform) method. The liner notes to their 1994 release (Example 13) provide basic information about their process.
[5.11] Fujieda’s subsequent cycle of pieces Patterns of Plants (1995–2011) exists for ensembles of Japanese instruments in various tuning systems. Sarah Cahill transcribed and performed some of the pieces for piano.(33) In the material accompanying the 2000 publication of piano transcriptions, Fujieda wrote of his desire to “hear [plants’] daily activities which are constantly changing as ‘their voices’” (Fujieda 2000, 2). However, it is clear that Fujieda’s biosignals are filtered and intermodulated quite freely through a complex mesh of different musical traditions including traditional Japanese chamber music, gamelan music and western postminimalism.(34) Fujieda writes:
One day when I was listening to a series of sounds generated from potentials of a plant, I intuitively felt that there must be beautiful melodies latent in them. I soon started to make a composition program based on plant potentials with [the] computer music program “MAX” so that I [could] find melodies in them.
As the title Patterns of Plants indicates, melodic patterns transcribed by [the] electric current of plants are incessantly proliferating. . . . The series of Patterns of Plants, which continues to develop in a recurrent process of combination and transformation, will not come to an end. (Fujieda 2009)
[5.12] This narrative alludes to a semiotic process we illustrated in Example 2. First, Fujieda listened to a “series of sounds generated from the potentials of a plant”—i.e., the unmitigated data that Dogane’s technology produced. Fujieda then had an intuition that these data contained something musical but he does not claim that the material itself was musical. Subsequently Fujieda exercised his own musical poiesis by writing a Max patch to find musical structures (“melodic patterns”) in the “incessantly proliferating” stream of data. He finally arranged these melodic patterns into compositions, completing the composer’s poietic process.
Audio Example 5. Fujieda: Pattern A from Patterns of Plants XI, Sarah Cahill, piano (Pinna Records 2, copyright © 2014, used with permission)
Example 14. Paradigmatic Analysis of Fujieda’s Pattern A from Patterns of Plants XI, piano version, top part only

(click to enlarge)
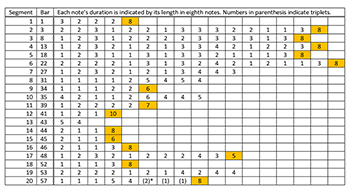
Example 17. Fujieda: Pattern A from Patterns of Plants XI: rhythmic analysis (segments that conclude with their longest values are highlighted in orange)
(click to enlarge)
[5.13] Pattern A from Patterns of Plants XI contains the fragmentary “melodic patterns” to which Fujieda alludes (Audio Example 5).(35) We parsed the top (“soprano”) melodic line of this piece into twenty phrases and performed a paradigmatic analysis, shown in Example 14. Our segmentation illustrates an enormous amount of repetition in the melodic contour and pitch profile. As indicated in Example 15, one four-element melodic contour segment, <-, +, -, ->, highlighted in bright orange, accounts for 56 of 160 contour changes. Together with the four-element contour segment that frequently follows it, <+, -, -, +>, shown in lighter orange, the two account for 96 of 160 contour changes. As Example 16 shows, the pitch content also contains frequent repetition, particularly the two incipit patterns <5, 4, 5, 4, 3> and <4, 3, 4, 3, 2> shown in bright orange and dark blue respectively (incidentally, the second incipit pattern is a literal transposition of the first, down one diatonic step.) However, the piece does not sound repetitive. How does Fujieda achieve variety? Fujieda structures the rhythm much differently than the pitch domain. As shown in Example 17, the durations (analyzed as eighth notes) are much more freely organized: it is not possible to find lengthy repeated patterns in the rhythmic dimension, though 15 of 20 phrases end in a relatively long duration, indicated in orange.(36)
Example 15. Fujieda: Pattern A from Patterns of Plants XI: contour analysis (different colors highlight repeating, or nearly repeating, patterns of contour)
(click to enlarge) | Example 16. Fujieda: Pattern A from Patterns of Plants XI: pitch analysis (different colors highlight repeating, or nearly repeating, patterns of pitches)
(click to enlarge) |
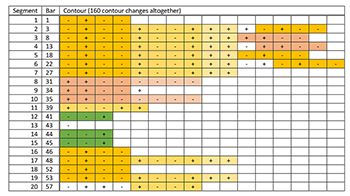
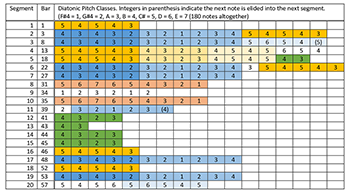
[5.14] Fujieda writes,
I extracted [a] variety of melodies from the sonic data of bio-electric fluctuations of the plants, which are, as it were, traces of their daily life. Then I bundled some of them as a “pattern.” (Fujieda 2000, 2)
These patterns emerge in our analysis as the repeating contours and pitch patterns. Given their stepwise simplicity, they are likely transcribed from short segments of plant data. It is clear that Fujieda’s content has been ordered, manipulated, and organized with a fairly strong hand, though his rhythmic durations are less predictable. Similar melodic and rhythmic structures occur in the other three contrapuntal voices. Here we make an inductive poietic claim based on analysis of the immanent structures of a written-out score, revealing Fujieda’s poietic task as twofold. First, Fujieda selected appropriate biodata segments from data produced by the Dogane’s Plantron, and then he arranged and transmuted these segments through the lens of several cultural traditions to produce a satisfactory score. Seen in this way, Fujieda’s technique of weaving together simple, repeating material into ever-changing textures, filtered through the vocabulary of different musical cultures, is remarkably complex.
6. Musical Style and EPM/S
[6.1] The processes biodata goes through in order to be transformed into sound imply no inherent musical idiom or style, scale, or system. As can be seen from our analysis, plant biodata must be mediated for it to conform to any recognizable musical genre. Nevertheless, artists frequently make choices to constrain data in some manner, positioning their work within a particular stylistic idiom. This gives us information about the many complex semiotic relationships in EPM/S. In this section, we interrogate the question of musical style in EPM/S and observe that composers generally locate “plant music” within two categories: ambient and postminimalist.
Ambient
[6.2] The origins of ambient music trace back to Erik Satie (Demers 2010, 116). Its history forms an arc spanning twentieth and twenty-first centuries, embracing work by composers including John Cage, La Monte Young, Suzanne Cianni, Meredith Monk, Karlheinz Stockhausen and many others (Toop 1995, 2018, 1–24). While bands such as Pink Floyd and Tangerine Dream are frequently cited as important influences, Brian Eno’s 1978 album “Music for Airports” is generally considered as an exemplar in the genre. Eno defined his view of ambient in opposition to what he called “canned music”:
Whereas the extant canned music companies proceed from the basis of regularizing environments by blanketing their acoustic and atmospheric idiosyncrasies, Ambient Music is intended to enhance these. . . . And whereas their intention is to “brighten” the environment by adding stimulus to it (thus supposedly alleviating the tedium of routine tasks and levelling out the natural ups and downs of the body rhythms) Ambient Music is intended to induce calm and a space to think. (Eno 1978)
[6.3] As evidence accumulates that plants themselves calm people’s physiology (Lee et al. 2015), the natural affinity EPM/S has with ambient music becomes more apparent. But herein lies one of the central challenges for musicians as they interpret biodata: how far should one go in mediating the data to produce a particular musical effect, and how much should a plant’s biological thumbprint (or “idiosyncracies”) remain? The “dial-it-in” approach of the PlantWave and Damanhour devices—which allow users to play music simply by selecting a preprogrammed patch—does not allow people to make these choices: instead, the software does all the work. This can “regularize” biodata into a homogenous smear of musical wash: precisely what Eno advocated against.
[6.4] Many EPM/S designers may gravitate towards the ambient genre because of how its adherents typically organize the time domain. Ambient music treats time in a way that seems to correspond to the view that the vegetal world exists outside of, or at least independent of, human time. Rupert Till clarifies this notion:
The musical style of ambient music affords a sense of kairos, or qualitative time, rather than chronos, quantitative time. A non-teleological musical approach allows pause, the creation of place, and the opportunity to slow the frantic pace of life for a moment. . . . Ambient music sets a context, a sense of place that is calming and serene, re- establishing a sense of home. It draws the listener inwards, stilling time and moving the deep listener into a sense of kairos, to a greater or lesser extent. (Till 2017)
[6.5] Leudar’s soundscapes in the rainforest were most obviously associated with ambient music’s stylistic markers, as they contained samples of natural sounds that were triggered by systems of communication native to the vegetal world itself, refusing to bow to human temporal exigencies. Data Garden’s hour-long tracks also exude this approach towards organizing time. In their music, bass notes seem to occur “when they do,” and one feels like having joined an ongoing process in media res; the fade-in and fade-out that bookend tracks correspond to the listener’s presumed entrance and exit. Here, the listener determines the frame, deemphasizing the composer’s authority to regulate the temporal extent of the work.
Postminimalism
[6.6] Michael Marder analyzes the temporal nature of plants from three perspectives: what he terms their “vegetal hetero-temporality of seasonal changes; the infinite temporality of growth . . . and the cyclical temporality of iteration, repetition and reproduction” (Marder 2013, 95). Whereas Marder’s second category is related to the kairotic view of time, seasonal changes and cyclical temporalities are connected to notions of repetition. What musical structures could transpose this element of phytotemporality into sound or music? According to Kyle Gann, postminimalist music often features phase-shifting of repetitive musical ideas, additive and subtractive processes, a “small, circumscribed set of materials,” as well as relatively consonant harmonies. He continues:
A postminimalist piece seems self-contained, not pointing outward; the references to other music sometimes contained therein are cut off from their source, preserved in abstract notes but not in emotional content, like a fly preserved in amber. . . . We are given only a circumscribed fragment of the musical universe to work with—and even within that truncated segment there is more going on than our ears and minds can account for. . . . The world, postminimalism seems to tell us, is understandable, but our perception is so limited that it (our perception) is easily overwhelmed by the interaction of even a few restricted elements and processes. (Gann 2013, 58)
[6.7] Data Garden’s music seems to fall into this aesthetic category too, perhaps because of the limited tonal means they employed and the repetitive patterns their performances hinged upon. Example 19 demonstrated that Fujieda’s piano piece works principally within a highly limited and repetitive melodic repertoire of only seven pitches; his three other contrapuntal parts are similarly constrained. Moreover, Fujieda incorporates subtractive and additive processes; Example 21 vividly shows how the duration of melodic structures is “pruned” through segments 11–13 only to “grow back” in subsequent segments. Finally, Fujieda’s integration of certain elements of Japanese court music, gamelan traditions, and western systems seems to provide the kind “circumscribed fragments” of musical worlds Gann writes about as hallmarks of a postminimalist style.
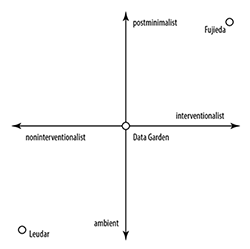
Example 18. Hypothetical scatter chart suggesting a correlation between the amount of intervention artists typically make interpreting biodata and the musical style that results
(click to enlarge)
[6.8] In the works we examined, we noticed that the amount of intervention a composer or sound designer made in interpreting biodata was related to the general style of the resulting artwork. In Example 18 we place the three works on a two-dimensional scatter plot where the x-axis represents the amount of intervention an artist employs in interpreting biodata, and the y-axis denotes the general stylistic character of the resulting project. Keeping in mind that assigning a style to a piece is often quite subjective, our representation nevertheless suggests a correlation. On one extreme lies Leudar, who discouraged the audience from touching plants, producing sound that aligns with the ambient genre most convincingly. In the middle lies the Data Garden Quartet, which neither encouraged nor discouraged listeners to touch their plants, but clearly used an array of quantizers to mediate data. Their music has qualities of both ambient and postminimalism. Fujieda extensively manipulated his biodata; in our view, his music shares the most affinity with postminimalism. Although our hypothesis is obviously preliminary on account of its small dataset, more work in this area could provide further insight.
7. EPM/S as a posthumanist/phytosemiotic avenue for encountering vegetal worlds
[7.1] Traversing the human and vegetal worlds invites many thorny questions, particularly when viewed in the context of western philosophical traditions. Assessing relationships between these domains has provoked several posthumanist authors to ask whether a significant reformulation, or even wholesale rejection of metaphysics, might be in order. Much of this work is indebted to Deleuze and Guattari (1987), who frequently employ metaphors (e.g., the rhizome) drawn from the phytosphere. More recently, Marder writes that “plants quietly subvert classical philosophical hierarchies” (Marder 2013, 53). The ecofeminist literature has advanced similar critiques (d’Eaubonne [1974] 2022; Ryan 2012; Plumwood 1993).
[7.2] Even from a very cursory perspective, humans and plants have deeply intertwined connections. Plants create the oxygen we breathe and feed us through the biomass they produce. We cannot survive without them, though they can (and have) existed without us. In a strictly materialistic economy, many people value domestic plants: houseplants enjoy a high level of prestige in a typical home’s “object ecology,” as rated alongside other objects (Csikszentmihalyi and Rochberg-Halton 1978). Jakob von Uexküll noted that plants create a “comfortable calm” that contrasts with the movement of animal life (von Uexküll 1922). Plants make us “feel” better: studies have shown that there are quantifiable physiological benefits when human subjects touch plants (Lee et al. 2015).(37) However, we will need more than this superficial analysis to recognize links between the human and vegetal domains in symbolic forms.
[7.3] A hundred years ago, Uexküll wrote of an organism’s Umwelt as its species-specific world of perception, arguing that life-forms are not simply the sum of their parts, but rather subjects with their own particular semiotic systems (von Uexküll 1922). If the phytosphere includes its own language of signals proper to itself, it is possible that it also communicates to us. Phytosemiotician Martin Krampen argues that plants portray forces in nature, reflected in the shape of their bodies (Krampen 1981, 207). They serve as meaningful signs to human endeavors because they are “living beings possessing features that evoke the attribution of meaning to a very considerable degree” (Krampen 1981, 207). Following upon the heels of this phytosemiotic critique, posthumanism positions the anthrosphere and phytosphere on a continuum, instead of at binary poles.
[7.4] The red arrows in Example 2 illustrate relationships between the anthrosphere and phytosphere which potentially influence outcomes of EPM/S. They cross over a liminal space where human and plant worlds meet. The act of surveying a foreign world from our own domestic gaze may invite us to consider whether a swerve towards ethnomusicology might sensitize us to the world of plants. This would be a step in the right direction, but ethnomusicologists typically work with human cultures: here, a more radical intervention seems to be in order. While it is tempting to think that elevating plants to the realm of “personhood” (by granting them some qualities we believe humans uniquely possess) may redress the effects of millennia of anthropocentric thinking, ascribing such qualities to plants burdens them with a notion of personhood to which they never consented. Marder enjoins us instead to approach the phytosphere by embracing the “non-cognitive, non-ideational, non-imagistic mode of thinking proper to plants”; to provocatively render human thinking “plant-like, altered by its encounter with the vegetal world” (Marder 2013, 10).
[7.5] Even so, Marder starts to sound like an ethnomusicologist when he advises us that if we wish to encounter plants on their own terms, we should preserve their alterity, avoid their objective description, and not presume an unconditional right to enter into the vegetal world. What grants us the right to have an encounter at all is the discovery of traces of ourselves in the plant domain, and vice-versa (Marder 2013, 9). Critiquing binary systems of thinking facilitates this encounter. Plants show us how tenuous such delineations are, even between our respective creative domains. Holly Watkins has written persuasively on the creative energy of plants (physis) and humans (techne), insisting that it is not possible to “enforce absolute distinctions between the two” (Watkins 2018, 8). Her work challenges the notion of human exceptionalism by showing the kinship between sound in the natural world and sound in human culture.
[7.6] In some EPM/S projects (particularly Leudar’s), human creators venture to encounter the phytosphere with minimal intervention. Although Data Garden’s lighthearted association of plants with performers in an imaginary ambient band may help some listeners better perceive musical forms by invoking familiar anthropocentric signposts, such markers unquestionably intrude into the vegetal world, which knows nothing of the peculiar structures of human music-making. Such insouciant experiments stray perilously close to a kind of techno-utopia that further colonizes the phytosphere. Future studies should consider the extent to which EPM/S projects participate in a resource-oriented extractive economy that enables exploitation of the natural world, or genuinely try to facilitate a deeper encounter with another realm of life.
[7.7] This reasoning leads us to close with an ethical question: how is it possible to have a just, equitable and musically satisfying interaction with the phytosphere? Plants do not normally participate in the human musical world on our terms, and we would be hard-pressed to describe what the sounds “proper to plants” might be. If one party seemingly has no stake in the outcome, how can true collaboration begin? To skeptics, EPM/S may seem like a triumphalist technological exercise that economically delivers an “incessantly proliferating” source of quasi-random information for sound designers to shape. Does EPM/S lend credibility to a false hope that we can access the vegetal through opaque layers of software and hardware, or is it a fertile field for mounting challenges to a reified metaphysics? Critical assessment of musical projects can provide much-needed perspectives to these questions and to the many others this article has posed.
[7.8] Although we remain skeptical of the ability to interact with the world of plants through EPM/S, the state of the art is still very new. Like Fujieda, who believed that bioelectric signals contained “traces of [plants’] daily life,” we remain optimistic and fascinated by the possibilities of EPM/S—not just by the way it promises to open up encounters with life-forms we depend on but still know so little about, but also by the way it challenges us to view the immense diversity of life with renewed wonder, awe, and empathy. This article has provided a broad but comprehensive footing on which further research can take root.
Paul V. Miller
Duquesne University
600 Forbes Avenue
Pittsburgh, PA 15282
millerp2@duq.edu
Christopher Cox
Duquesne University
600 Forbes Avenue
Pittsburgh, PA 15282
cmcox@gmail.com
Works Cited
Affifi, Ramsey. 2013. “Learning Plants: Semiosis Between the Parts and the Whole.” Biosemiotics 6 (3): 547–59. https://doi.org/10.1007/s12304-013-9164-x.
Agawu, Kofi. 1987. “The Rhythmic Structure of West African Music.” The Journal of Musicology 5 (3): 400–418. https://doi.org/10.2307/763699.
Alpi, Amedeo, et al. 2007. “Plant Neurobiology: No Brain, No Gain?” Trends in Plant Science 12 (4): 135–36. https://doi.org/10.1016/j.tplants.2007.03.002.
Baluška, František, and Stefano Mancuso, eds. 2009. Signaling in Plants. Springer.
Baluška, František, Stefano Mancuso, and Dieter Volkman, eds. 2006. Communication in Plants: Neuronal Aspects of Plant Life. Springer.
Beloff, Laura. 2020. “The Hearing Test: Evidence of a Vegetal Entity.” Leonardo Music Journal (30): 85–89. https://doi.org/10.1162/lmj_a_01097.
Body, Mélanie J. A., et al. 2019. “Caterpillar Chewing Vibrations Cause Changes in Plant Hormones and Volatile Emissions in Arabidopsis thaliana.” Frontiers in Plant Science 10: 810. https://doi.org/10.3389/fpls.2019.00810.
Boyno, Gökhan, and Semra Demir. 2022. “Plant-Mnycorrhiza Communication and Mycorrhizae in Inter-plant Communication.” Symbiosis 86: 155–68. https://doi.org/10.1007/s13199-022-00837-0.
Brenner, Eric, et. al. 2007. “Response to Alpi et al.: Plant Neurobiology: The Gain is More than the Name.” Trends in Plant Science 12 (7): 285–86. https://doi.org/ 10.1016/j.tplants.2007.06.005.
Burdon-Sanderson, John Scott. 1873. “Note on the Electrical Phenomena which Accompany Irritation of the Leaf of Dionaea muscipula.” Proceedings of the Royal Society of London 21: 495–96. https://doi.org/10.1098/rspl.1872.0092.
Canales, Javier, Carlos Henriquez-Valencia, and Sebastian Brauchi. 2018. “The Integration of Electrical Signals Originating in the Root of Vascular Plants.” Frontiers in Plant Science 8: 1–15. https://doi.org/10.3389/fpls.2017.02173.
Chamovitz, Daniel. 2012. What a Plant Knows: A Field Guide to the Senses. Scientific American; Farrar, Straus and Giroux.
Csikszentmihalyi, Mihaly, and Eugene Rochberg-Halton. 1978. “Reflections on Materialism: People and Things.” The University of Chicago Magazine 70 (3): 7–15. https://campub.lib.uchicago.edu/view/?docId=mvol-0002-0070-0003#page/1/mode/1up.
Darwin, Charles. 1898. The Power of Movement in Plants. D. Appleton and Co. https://doi.org/10.5962/bhl.title.56998.
d’Eaubonne, Francoise. (1974) 2022. Feminism or Death: How the Women's Movement Can Save the Planet. Translated by Ruth Hottell. Verso.
Deleuze, Gilles, and Félix Guattari. 1987. A Thousand Plateaus: Capitalism and Schizophrenia. University of Minnesota Press.
Demers, Joanna. 2010. Listening Through the Noise: The Aesthetics of Experimental Electronic Music. Oxford University Press.
Duursma, Remko A., et. al. 2019. “On the Minimum Leaf Conductance: Its Role in Models of Plant Water Use, and Ecological and Environmental Controls.” New Phytologist 221 (2): 603–705. https://doi.org/10.1111/nph.15395.
Fromm, Jörg, and Silke Lautner. 2006. “Electrical Signals and their Physiological Significance in Plants.” Plant, Cell and Environment 30 (3): 249–57. https://doi.org/10.1111/j.1365-3040.2006.01614.x.
Fujieda, Mamoru. 2000. Patterns of Plants. Ongaku No Tamo Sha.
—————. 2009. “Tuning of ‘Patterns of Plants’.”� In Arcana IV: Musicians on Music, ed. John Zorn, 140–50. Hips Road.
Galston, Arthur W., and Clifford L. Slayman. 1979. “The Not-So-Secret Life of Plants: In Which the Historical and Experimental Myths About Emotional Communication Between Animal and Vegetable are Put to Rest.” American Scientist 67 (3): 337–44. http://www.jstor.org/stable/27849226.
Gann, Kyle. 2013. “A Technically Definable Stream of Postminimalism, Its Characteristics and Its Meanings.” In The Ashgate Research Companion to Minimalist and Postminimalist Music, ed. Keith Potter and Kyle Gann. Routledge, 39–60.
Grond, Florian, and Thomas Hermann. 2012. “Aesthetic Strategies in Sonification.” AI and Society 27 (2): 213–22. https://doi.org/10.1007/s00146-011-0341-7.
Johns, Sarah, Takuma Hagihara, Masatsugu Toyoto, and Simon Gilroy. 2021. “The Fast and the Furious: Rapid Long-Range Signaling in Plants.” Plant Physiology 185 (3): 694–706. https://doi.org/10.1093/plphys/kiaa098.
Jung, Jihye, et al. 2018. “Beyond Chemical Triggers: Evidence for Sound-Evoked Physiological Reactions in Plants.” Frontiers in Plant Science 9 (25). https://doi.org/10.3389/fpls.2018.00025.
Khait, I., et al. 2019. “Plants Emit Informative Airborne Sounds Under Stress.” bioRxiv (December). https://doi.org/10.1101/507590.
Krampen, Martin. 1981. “Phytosemiotics.” Semiotica 36 (3–4): 187–209. https://doi.org/10.1515/semi.1981.36.3-4.187.
l’Anson, Mileece. 2019. “Form, Chaos, and the Nuance of Beauty.” In The Oxford Handbook of Algorithmic Music, ed. Roger T. Dean and Alex McLean. Oxford University Press. https://doi.org/10.1093/oxfordhb/9780190226992.013.33.
Larimore, Franklin C. 1984. T.E.N.S. Electrode. US Patent 4,458,696, issued July 10.
Larsson, Naomi. 2022. “Button Pushers: The Artists Making Music from Mushrooms.” The Guardian, May 13, 2022.
Lee, Min-Sun, et al. 2015. “Interaction with Indoor Plants May Reduce Psychological and Physiological Stress by Suppressing Autonomic Nervous System Activity in Young Adults: A Randomized Crossover Study.” Journal of Physiological Anthropology 34 (21). https://doi.org/10.1186/s40101-015-0060-8.
Leudar, Augustine. 2014. “Invisible Forest.” Plasticitiés Sciences Arts 34.
Leudar, Augustine. 2018. “Surrounded: A Series of Sound Installations That Combine Plant Electrophysiology and 3D Sonic Art.” Leonardo 51 (5): 517–23. https://doi.org/10.1162/leon_a_01338.
Macedo, Francynês da Conceiçao Oliveira, et. al. 2021. “Equipment and Protocol for Measurement of Extracellular Electrical Signals, Gas Exchange and Turgor Pressure in Plants.” MethodsX 8 (101214). https://doi.org/10.1016/j.mex.2021.101214.
Marder, Michael. 2013. Plant-Thinking: A Philosophy of Vegetal Life. Columbia University Press.
Miller, Paul, and Brian Riordan. 2019. “Can A Leaf Make Music? Techniques and Aesthetics of Plant-Generated Control Voltage in Electronic Music.” Poster session presentation at the National Meeting of the Society for Music Theory, November 8, Columbus, Ohio. http://theoryofpaul.net/plants/poster-07.pdf.
Nattiez, Jean-Jacques. 1990. Music and Discourse: Toward a Semiology of Music. Translated by Carolyn Abbate. Princeton University Press.
Plumwood, Val. 1993. Feminism and the Mastery of Nature. Routledge.
Pollan, Michael. 2013. “Intelligent Plants.” The New Yorker, December 23, 2013.
Ryan, John Charles. 2012. “Passive Flora? Reconsidering Nature’s Agency through Human-Plant Studies (HPS).” Societies 2 (3): 101–21. https://doi.org/10.3390/soc2030101.
Shabala, Sergey, and Stefano Mancuso, eds. 2007. Rhythms in Plants: Phenomenology, Mechanisms and Adaptive Significance. Springer.
Simard, Suzanne W. 2018. “Mycorrhizal Networks Facilitate Tree Communication, Learning, and Memory.” In Memory and Learning in Plants: Signaling and Communication in Plants, ed. František Baluška, Monica Gagliano, and Guenther Witzany, 191–213. Springer. https://doi.org/10.1007/978-3-319-75596-0_10.
Stankovic, Bratislav, Daniel LK. Witters, Tadeusz Zawadski, and Eric Davies. 1998. “Action Potentials and Variation Potentials in Sunflower: An Analysis of their Relationships and Distinguishing Characteristics.” Physiologia Plantarium 103 (1): 51–58. https://doi.org/10.1034/j.1399-3054.1998.1030107.x.
Stockhausen, Karlheinz. 1978. Texte zur Musik. Vol. 4. DuMont.
—————. 2009. Kompositorische Grundlagen Neuer Musik: Sechs Seminare für die Darmstädter Ferienkurse 1970. Stockhausen Stiftung für Musik.
Till, Rupert. 2017. “Ambient Music and Religion.” Bloomsbury Handbook of Religion and Popular Music, ed. Christopher Partridge and Marcus Moberg, 327–37. Bloomsbury. https://doi.org/10.5040/9781474237369.0038.
Tipp, Devon Osamu. 2023. “Mamoru Fujieda’s Plant Language: Hybrid Approaches to Composing for Koto.” PhD diss., University of Pittsburgh.
Tompkins, Peter, and Christopher Bird. 1973. The Secret Life of Plants. HarperCollins.
Toop, David. 1995, 2018. Ocean of Sound: Ambient Sound and Radical Listening in the Age of Communication. Serpent's Tail.
Trewavas, Anthony. 2007. “Response to Alpi et al.: Plant neurobiology – All Metaphors Have Value.” Trends in Plant Science 12 (6): 231–33. https://doi.org/10.1016/j.tplants.2007.04.006.
Tsadka, Maayan. 2020. “A Field Guide to Sonic Botany: Thoughts about Eco-Composition.” Tempo 75 (295): 31–44. https://doi.org/10.1017/S0040298220000662.
Uexküll, Jakob von. 1922. “Wie sehen wir die Natur und wie sieht sie sich selber?” Die Naturwissenschaft 10: 296–301. https://doi.org/10.1007/BF01565504.
Watkins, Holly. 2018. Musical Vitalities: Ventures in a Biotic Aesthetics of Music. The Chicago University Press.
Xie, Jinjin, Yanyou Wu, Deke Xing, Zhongying Li, Tian Chen, Rongrong Duan, and Xiaoxing Zhu. 2021. “A Comparative Study on the Circadian Rhythm of the Electrical Signals of Broussonetia papyrifera and Morus alba.” Plant Signaling and Behavior 16 (11): 1950899. https://doi.org/10.1080/15592324.2021.1950899.
Discography
Discography
Data Garden Quartet. 2012. Live at the Philadelphia Museum of Art. https://datagarden.bandcamp.com/album/quartet-live-at-the-philadelphia-museum-of-art.
Eno, Brian. 1978. Ambient 1: Music for Airports. Polydor Records 7908.
Fujieda, Mamoru, and Yuji Dogane. 1994. Ecological Plantron. K-Planning, KPCD-01, LMCD-1420, EM Records EM 1202 (LP). https://emrecords.bandcamp.com/album/ecological-plantron.
Fujieda, Mamoru, and Yuji Dogane. 1997. Patterns of Plants. Tzadik TZ 7025.
Fujieda, Mamoru, and Yuji Dogane. 2014. Patterns of Plants, Sarah Cahill, piano. Pinna Records 2.
Footnotes
1. This definition excludes compositions that involve mechanically generating sound with plants. John Cage’s amplified cactus pieces (Child of Tree, 1975 and Branches, 1976) lie beyond our scope because they hinge on a performer’s physical interaction with the plant itself. The phenomenon of the “amplified cactus” has reverberated in the works of Paul Rudy (Grandchild of Tree, 2000) and Mark Andre (. . .zu Staub. . ., 2017). Other representative works outside the framework of EPM/S include Karlheinz Stockhausen’s Herbstmusik, which is constructed of sounds produced by sticks, wheat, and leaves (Stockhausen 1978, 246–47) and Maayan Tsadka’s Sonic Botany (Tsadka 2020).
Return to text
2. Nattiez defined the symbolic form as “a sign, or collection of signs, to which an infinite complex of interpretants is linked” (Nattiez 1990, 8).
Return to text
3. Plants do emit ultrasonic sounds in the range of 20–100kHz (Khait et al. 2019). Artists have already begun to use these sounds creatively (Beloff 2020). In addition to their actual production of sound, there is robust evidence to show that plants are sensitive to it (Body et al. 2019). This is surprising because plants do not possess specialized auditory sensory organs; nevertheless, they exhibit significant genetic transcriptional and hormonal changes when exposed to the minute rarefactive and compressive physical forces of sound pressure waves (Jung et al. 2018). Exploring plants’ sonic feedback cycle has formed the basis of Tarun Nayer’s work and may hold promise for future exploration (Larsson 2022).
Return to text
4. The field of plant “neurobiology” and the meaning of the word “neuron” prompted a widespread critical reassessment of the term (Brenner et al. 2007; Trewavas 2007; Alpi et al. 2007).
Return to text
5. In experimental data, resistance and impedance peak at about 2:00 pm while capacitance peaks at 2:00 am. This is not surprising because the resistance between conductors is inversely proportional to their capacitance.
Return to text
6. For a comprehensive overview from a lay perspective, see Chamovitz 2012.
Return to text
7. http://www.linv.org/about-us/, accessed 1 August 2023.
Return to text
8. Common plant species are also determined by their availability in greenhouses and home improvement stores, as most artists do not have access to specialized species. Some typical examples include Crassula ovata (jade plant), Dracena fragrans (cornstalk plant), Sansevieria trifasciata (snake plant), Codiaeum (croton), Monstera deliciosa (Swiss cheese plant), Ficus and Zamioculcas zamiifolia (ZZ plant).
Return to text
9. This definition is accurate but oversimplifies the complexity of the MIDI standard. For more information about MIDI, see https://www.midi.org/specifications (accessed 24 July 2023).
Return to text
10. While writing this article, we learned of yet another commercial biosonification device, similar to PlantWave but even more simple called “PlantsPlay.” See https://plantsplay.com, accessed 24 July 2023.
Return to text
11. https://github.com/electricityforprogress/MIDIsprout, accessed 24 July 2023.
Return to text
12. It is easy to demonstrate that this is the operational principle by inserting a variable resistor (such as a potentiometer) between the two TENS electrodes. When the resistance is changed, MIDISprout generates a rapid, wide pitch range of MIDI notes; holding the trimmer stable results in a dramatically less frequent, narrow range.
Return to text
13. Interrupts measure the 555 signal at a variable sample rate that is at minimum 2 microseconds (2 MHz).
Return to text
14. The principle resonates with certain historical techniques of serial composition. An important aspect of Stockhausen’s serial technique in Kontakte (1958–60) hinges on the amount of change in integer magnitudes derived from serial matrices, not the magnitudes themselves. Stockhausen employed “Reihe der Veränderungsgrad” (“series of degrees of change”) to inspire subjective compositional choices regarding spatialization, instrumentation, form, tempo, register, and dynamics (Stockhausen 2009, 232–38).
Return to text
15. For more on this system, see https://www.adafruit.com (accessed 1 August 2023).
Return to text
16. https://damanhur.org, accessed 24 July 2023.
Return to text
17. This is based on an SAM2695, which is essentially a “synthesizer on a chip”.
Return to text
18. The LabJack is a data acquisition device used in some laboratory environments. See https://labjack.com/products/u3, accessed 24 July 2023.
Return to text
19. Dieter Doepfer specified the Eurorack format in 1996. The technical specifications are available at https://doepfer.de/a100e.htm, accessed 24 July 2023.
Return to text
20. https://www.instruomodular.com, accessed 24 July 2023.
Return to text
21. The Damanhour device contains a much larger ADM3260 isolator as well, presumably for the same purpose.
Return to text
22. We set the Damanhour to “chromatic” mode, where MIDI pitches are quantized to an equal-tempered western scale.
Return to text
23. It is possible that the maximum threshold setting on Scion’s sensitivity dial is set too low. This would be easy to fix in a Firmware upgrade.
Return to text
24. Note that this data is control voltage, not MIDI data or an audio signal (even though it is sampled at audio rate).
Return to text
25. The term “transient” here is used in its acoustic context, i.e., a short, high-amplitude component that frequently occurs at the beginning of a musical sound, noise or vocalization.
Return to text
26. https://vimeo.com/63343503, accessed 1 August 2023.
Return to text
27. A short documentary illustrating Leudar’s installation work can be found at https://www.youtube.com/watch?v=nhomVIY9CQY, accessed 24 July 2023.
Return to text
28. Recent research has found that plants communicate with each other via sophisticated subterranean mycorrhizal networks (Simard 2018; Boyno and Demir 2022).
Return to text
29. Leudar used ambisonic panners developed for Max/MSP at the Institute for Computer Music and Sound Technology (ICST, Zurich University of the Arts). See https://www.zhdk.ch/en/research/icst/software-downloads-5379/downloads-ambisonics-externals-for-maxmsp-5381, accessed 1 August 2023.
Return to text
30. https://datagarden.bandcamp.com/album/quartet-live-at-the-philadelphia-museum-of-art, accessed 24 July 2023. A short video is also available at https://www.youtube.com/watch?v=ekV1ngd_9GY.
Return to text
31. Fujieda studied composition with Morton Feldman and others at the University of California, Santa Cruz in the 1980s. His compositional influences include Harry Partch and Lou Harrison.
Return to text
32. This, too, is available on Bandcamp: https://emrecords.bandcamp.com/album/ecological-plantron, accessed 24 July 2023.
Return to text
33. Pinna Records 2, http://www.pinnarecords.com/products/530282-mamoru-fujieda-patterns-of-plants-sarah-cahill-piano, accessed 1 August 2023.
Return to text
34. Devon Osamu Tipp has attempted the Herculean task of unpacking the various cultural influences. After cataloging many Japanese and Indonesian traits, Tipp concludes that Fujieda’s adoption of the western equal tempered system in some Collections amounts to a jarring cultural dissonance (Tipp 2023, 52).
Return to text
35. Although the piece was originally known as Patterns of Plants 3, Pattern A, it was published for the piano as Patterns of Plants XI, Pattern A (Fujieda 2000). This is the version we analyze. In the original, Fujieda employs three microtonally tuned kotos. According to Tipp, this piece functions like a section of a typical four-movement Japanese gagaku suite. Its melodic structures are reminiscent of techniques in the jiuta-sokyoku tradition of Japanese court chamber music. Tipp notes that Pattern A references elements from Indonesian gamelan music as well.
Return to text
36. Fujieda’s proclivity to conclude phrases with a long note in this piece (and his few other gamelan-inspired works) evokes the “gong chord� common in Indonesian gamelan music.
Return to text
37. The houseplant market exploded during the COVID-19 pandemic. Its market value in the United States alone was a staggering $18 billion in 2021. https://www.prnewswire.com/news-releases/indoor-plants-market-to-register-promising-growth-of-usd-26-23-billion-by-2029--size-share-growth-opportunities-trade-insights-and-competitive-landscape-301655694.html, accessed 25 July 2023.
Return to text
Copyright Statement
Copyright © 2024 by the Society for Music Theory. All rights reserved.
[1] Copyrights for individual items published in Music Theory Online (MTO) are held by their authors. Items appearing in MTO may be saved and stored in electronic or paper form, and may be shared among individuals for purposes of scholarly research or discussion, but may not be republished in any form, electronic or print, without prior, written permission from the author(s), and advance notification of the editors of MTO.
[2] Any redistributed form of items published in MTO must include the following information in a form appropriate to the medium in which the items are to appear:
This item appeared in Music Theory Online in Volume 30, Issue 1 in March 2024. It was authored by Paul V. Miller (millerp2@duq.edu) and Christopher Cox (cmcox@gmail.com), with whose written permission it is reprinted here.
[3] Libraries may archive issues of MTO in electronic or paper form for public access so long as each issue is stored in its entirety, and no access fee is charged. Exceptions to these requirements must be approved in writing by the editors of MTO, who will act in accordance with the decisions of the Society for Music Theory.
This document and all portions thereof are protected by U.S. and international copyright laws. Material contained herein may be copied and/or distributed for research purposes only.

Prepared by Lauren Irschick, Editorial Assistant
Number of visits:
30293